우리 연구실은 크게 애기장대를 대상으로 (1) RNA 생물학, (2) 식물 발달생물학, (3) 식물 스트레스 생물학을 연구하고 있습니다. 이를 위해 돌연변이체를 활용한 분자유전학, 분자생물학, 그리고 세포생화학적 기법을 사용하고 있습니다.
또, 유채를 대상으로 애기장대의 연구결과를 활발하게 적용하여 응용하고자 합니다. 유채는 애기장대와 같은 십자화과(배추과)에 속하기때문에 애기장대의 결과가 유채에 잘 반영될 수 있을 것입니다.아래 연구의 설명에서 유전자 이름이 이탤릭 대문자(예, STA1)로 쓰인 것은 정상 유전자를 의미합니다. 이탤릭 소문자(예, sta1)로 쓰인 것은 돌연변이 유전자 또는 돌연변이 유전자를 가진 돌연변이 식물체를 의미합니다.
(1) RNA 생물학: RNA 대사, 비암호화 RNA 생성, DNA 메틸화, 온도스트레스 특이적 반응
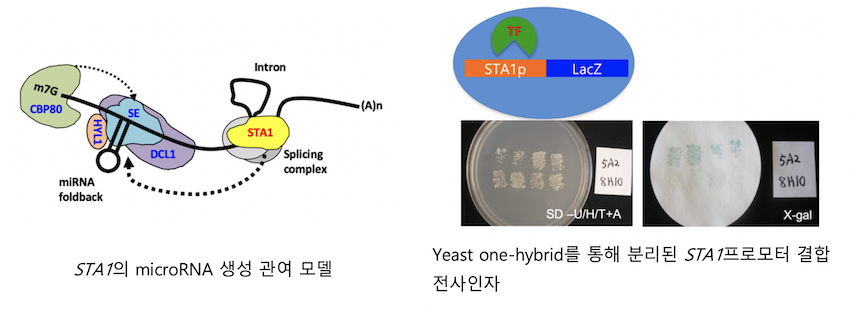
애기장대 STABILIZED1 (STA1) 유전자는pre-mRNA splicing factor인 PRP6 유사단백질을 암호화하고 있고 mRNA splicing과 stability에 관여하고 있습니다. 우리는 STA1 유전자가 splicing 기능을 통해서 small noncoding RNA (소형 비암호화 RNA)인 microRNA 생성에도 관여함을 밝혔습니다. 또 한편, STA1이 RNA-directed DNA methylation (RdDM) 에도 관여함이 밝혀져 있습니다.
이에 우리는 이 연구를 확장하기 위해서 STA1을 중심으로 하는, RNA 대사, 비암호화 RNA 생성, RdDM 기작을 중점적으로 연구하고 있습니다. 이를 위해 sta1-1 돌연변이체 결함이 증가된 enhancer 돌연변이체와 결함이 감소된 suppressor 돌연변이체를 확보하였고, 이들에서 STA1과 유전학적으로 상호작용하는 유전자를 찾고 있습니다.
한편, STA1이 splicing factor이기 때문에, 세포 내에서 늘 필요할 것 같음에도, 온도 스트레스에 의해 그 발현이 유도됩니다. 이것은 항상 기능할 것 같은 STA1이 온도스트레스에서 특이적인 기능을 가진다는 것을 의미합니다. 이에, 온도스트레스에서 STA1 발현을 조절하는 유전자를 찾고 있습니다. Yeast one-hybrid 기법을 이용해서 STA1 프로모터에 결합하는 두 전사인자를 분리하였고 이에 대한 특징과 기능을 분석하고 있습니다. 또한 STA1 프로모터에 luciferase를 결합한 리포터 유전자를 가지는 애기장대를 이용하여STA1 발현이 변화한 돌연변이체를 분리하였고 이 돌연변이체의 특징과 그 돌연변이를 일으키는 유전자, 즉 STA1 발현을 조절하는 유전자를 찾고 있습니다.
(2) 식물 발달프로그램: SGAL1유전자의 기능
우리는 식물 형태 결함 돌연변이체인 small and glassy leaf 1 (sagl1) 애기장대 돌연변이체를 발굴하였습니다. 이름이 말해주듯이 이 돌연변이체는 작고 반짝이는,그리고 두꺼운 잎을 가지고 있습니다. 잎이 반짝이는 이유는 이 sagl1 돌연변이체의 잎 표면층에 더 많은 왁스를 축적하고 있기 때문 입니다. 또, 한편 sagl1 돌연변이체는 붉은 색소인 anthocyanin을 많이 축적합니다. 우리는 최근 이 sagl1 돌연변이체 내의 어떤 유전자가 결함이 생겨서 이러한 표현형을 보이게 하는 지를 알아 냈습니다. SAGL1 유전자의 기능 연구를 통해 잎의 왁스 축적과 anthocyanin 축적이 이 돌연변이체에서 많이 일어나는 이유를 밝혔습니다.
한편, sagl1 돌연변이체에서 식물의 줄기세포의 하나라고 할 수 있는 형성층이 증가한 것을 관찰하여,식물 형성층 줄기세포에서의 SAGL1 유전자 기능 연구에도 초점을 맞추고 있습니다.

(3) 식물 스트레스 생물학: 스트레스 신호전달과 다양한 대응 기작, 유전자가위의 활용
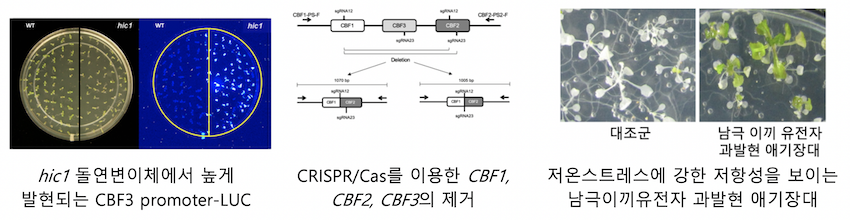
애기장대에서 C-repeat binding factor (CBF)는 식물 저온 스트레스 신호전달에서 중요한 전사인자입니다. 애기장대에는 저온 스트레스 신호전달에 관여하는 CBF 전사인자가 3개 존재하고 각각 CBF1, CBF2, CBF3라고 합니다. 우리는 CBF3 프로모터에 luciferase 유전자를 결합한 리포터 시스템을 이용해서 CBF3 발현이 변화한 돌연변이체를 찾아냈습니다. 이 중 하나인 highly induced CBF3 expression1 (hic1)은 저온에서 정상체보다 더 높은 CBF 유전자 발현을 보입니다. 최근 우리는 이 hic1돌연변이체 결함이 어떤 유전자에 있기 때문인지를 알아냈습니다. 즉, CBF유전자 발현에 중요한 유전자를 찾은 셈입니다. 우리는 이 HIC1유전자의 기능을 분석 중에 있습니다.
또한 우리는 유전자가위인 CRISPR/Cas 시스템을 이용하여 애기장대의 CBF1, CBF2, CBF3유전자를 모두 제거하는 데 성공했습니다. 특히, 애기장대의 일종의 아종(accession 또는 ecotype)이라고 할 수 있는 Columbia-0, C24, Landsberg erecta에서 모두 CBF1, CBF2, CBF3 유전자를 삭제하여 각 다른 애기장대 accession에서 저온신호전달을 연구할 수 있는 토대를 만들었습니다.
아울러, 극지연구소와 공동연구로, 남극에 존재하는 이끼에서 분리한 유전자를 애기장대에 과발현시켜 더 강한 스트레스 저항성을 갖게 하는 유전자를 찾고 있습니다. 이미 애기장대에서 더 강한 스트레스 저항성을 가지게 하는 극지 이끼 유전자를 수 종을 확보하였습니다. 우리는 이와 같은 극지연구소와의 공동연구를 통해서 극지 유전자원의 활용도를 높일 수 있을 것으로 기대합니다. 우리 연구실은 국내에서 CRISPR/Cas 유전자 가위를 식물에 적용한 선두 그룹으로 이렇게 발굴된 극지 이끼 유전자의 특징을 애기장대와 유채에 CRISPR/Cas기술로 도입하여 스트레스 저항성 식물 연구 및 활용에 적극 활용하고 있습니다.